
Raman Spectroscopy of Oral Candida Species: Molecular-Scale Analyses, Chemometrics, and Barcode Identification

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
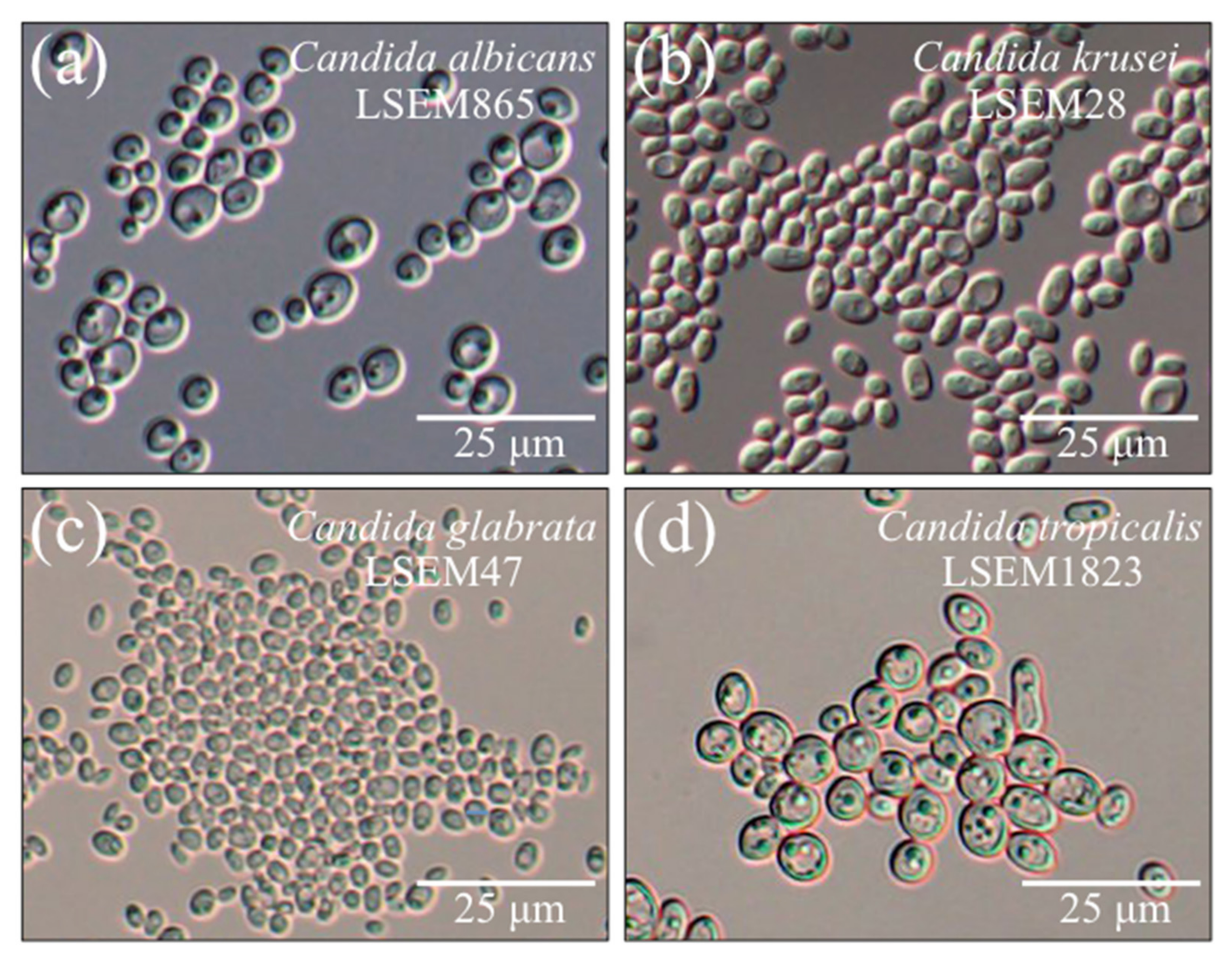
2.1. Morphological Observations of the Studied Species
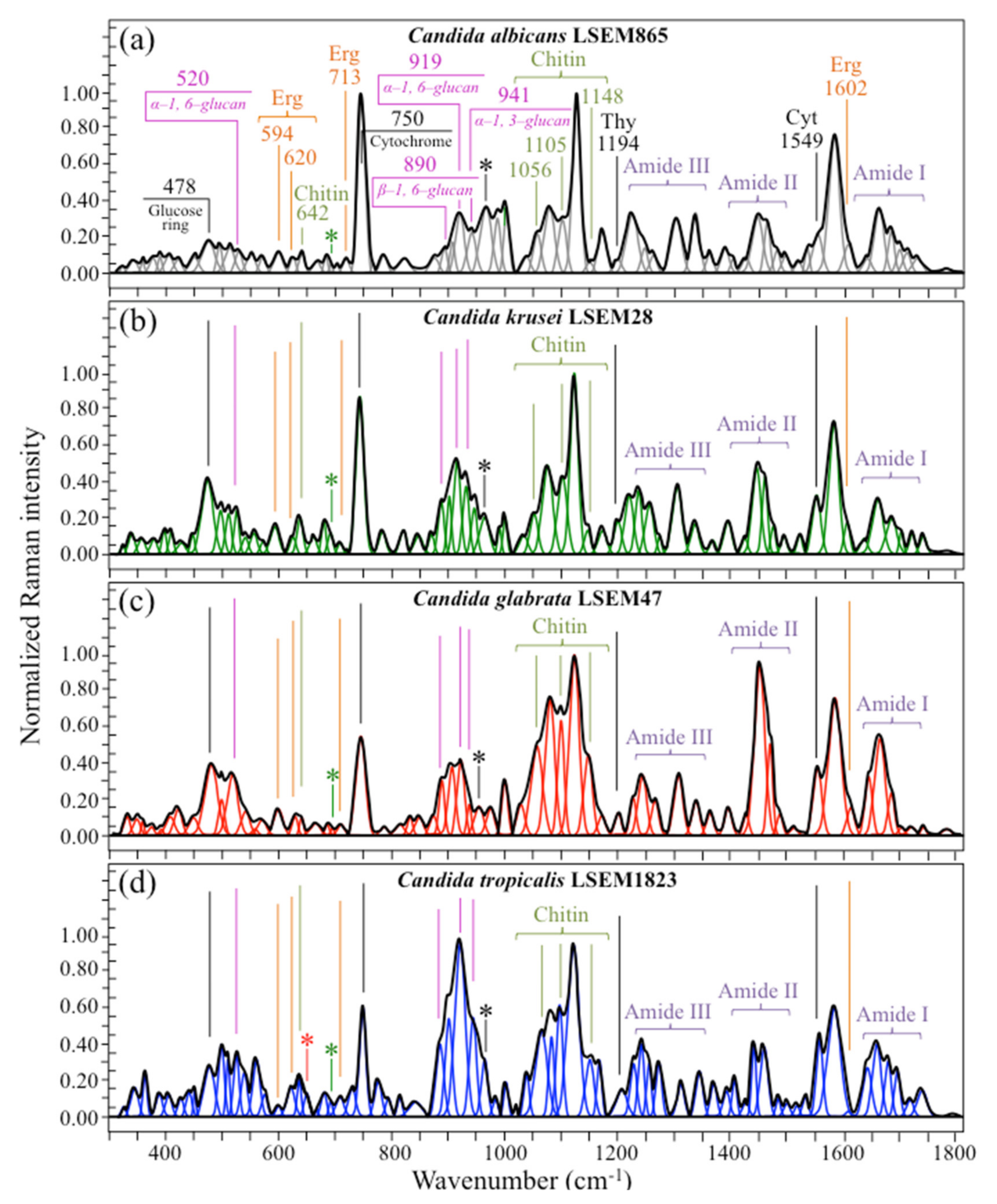
2.2. Raman Analyses of Different Candida Species
- (i)
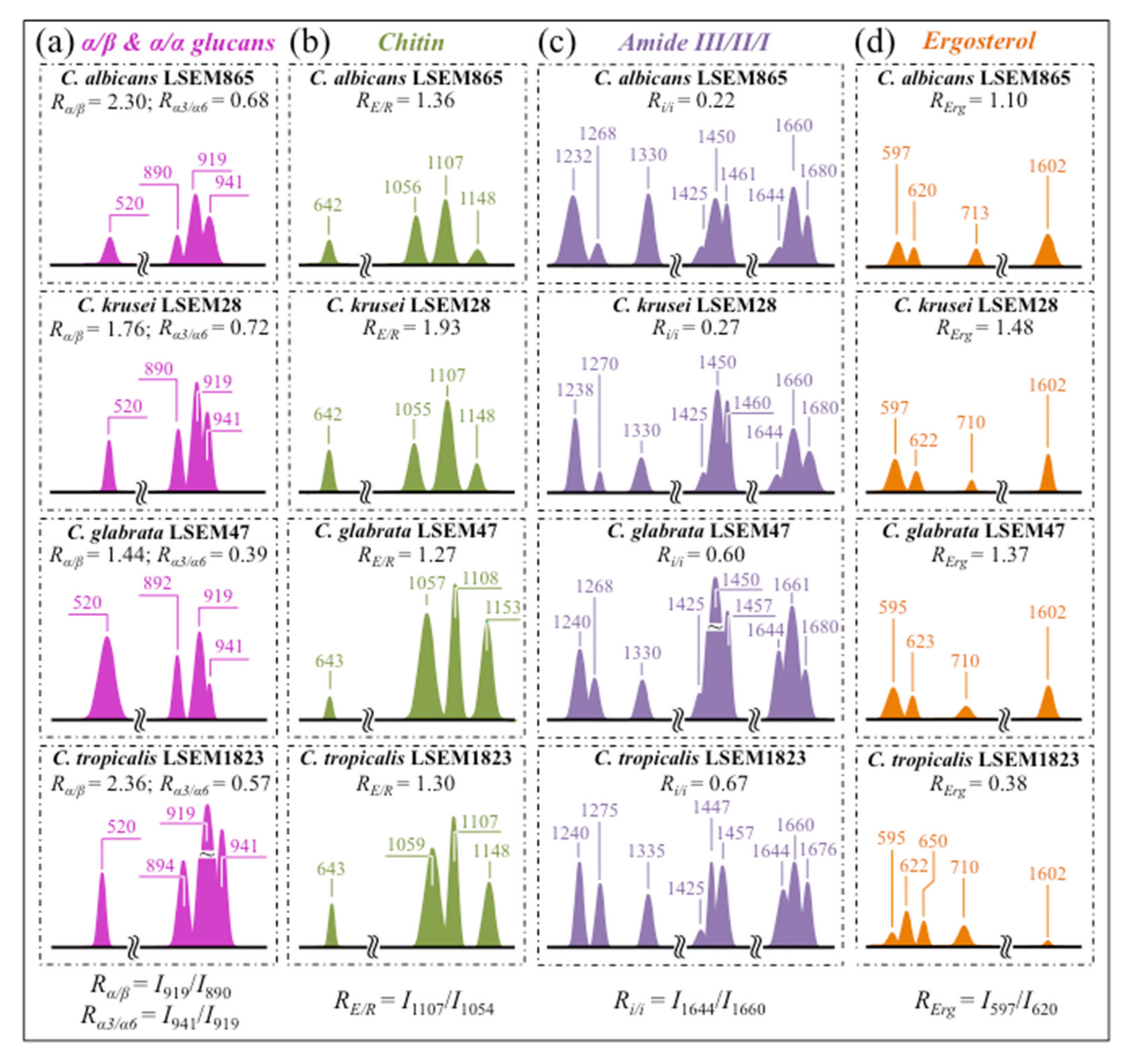
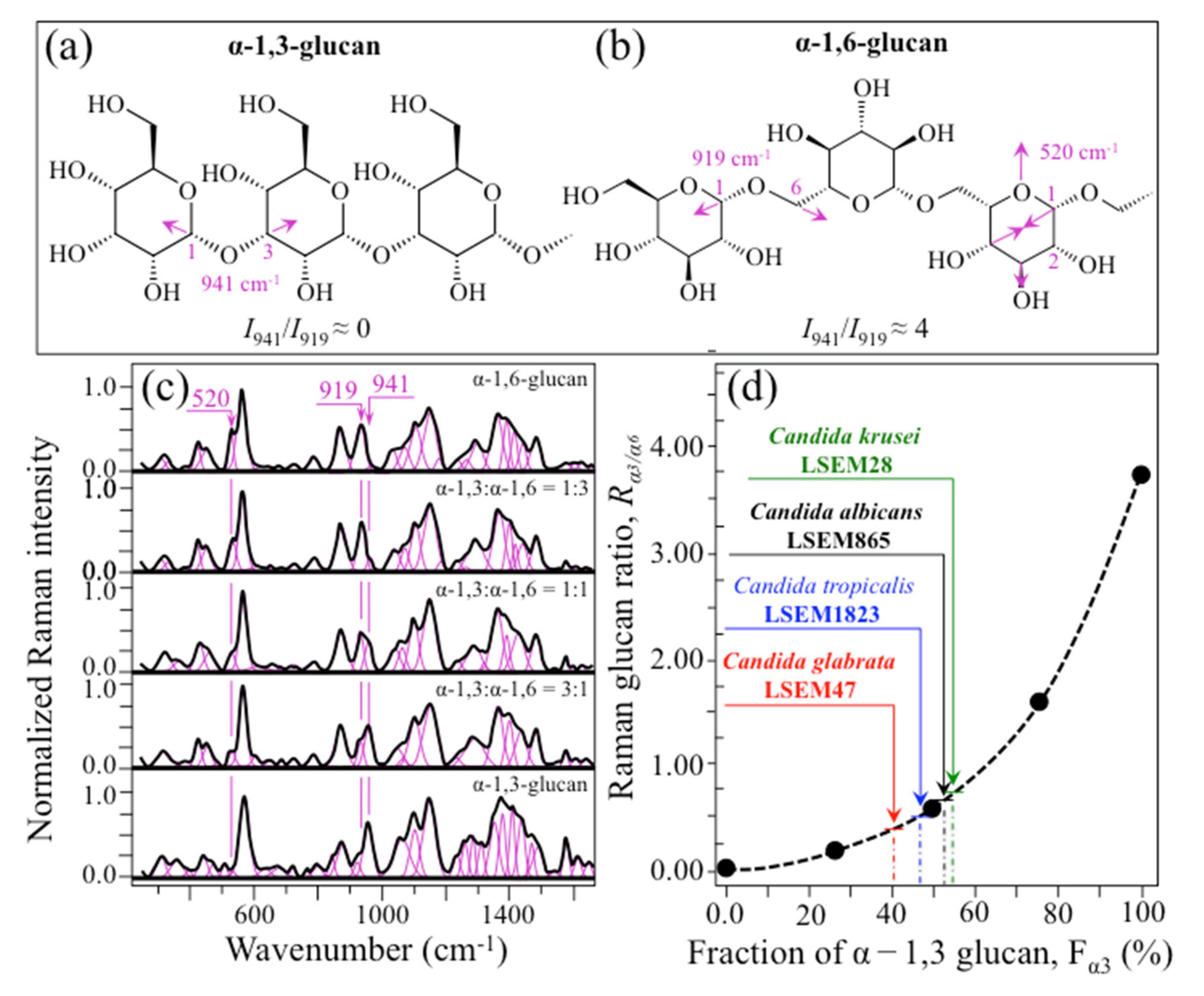
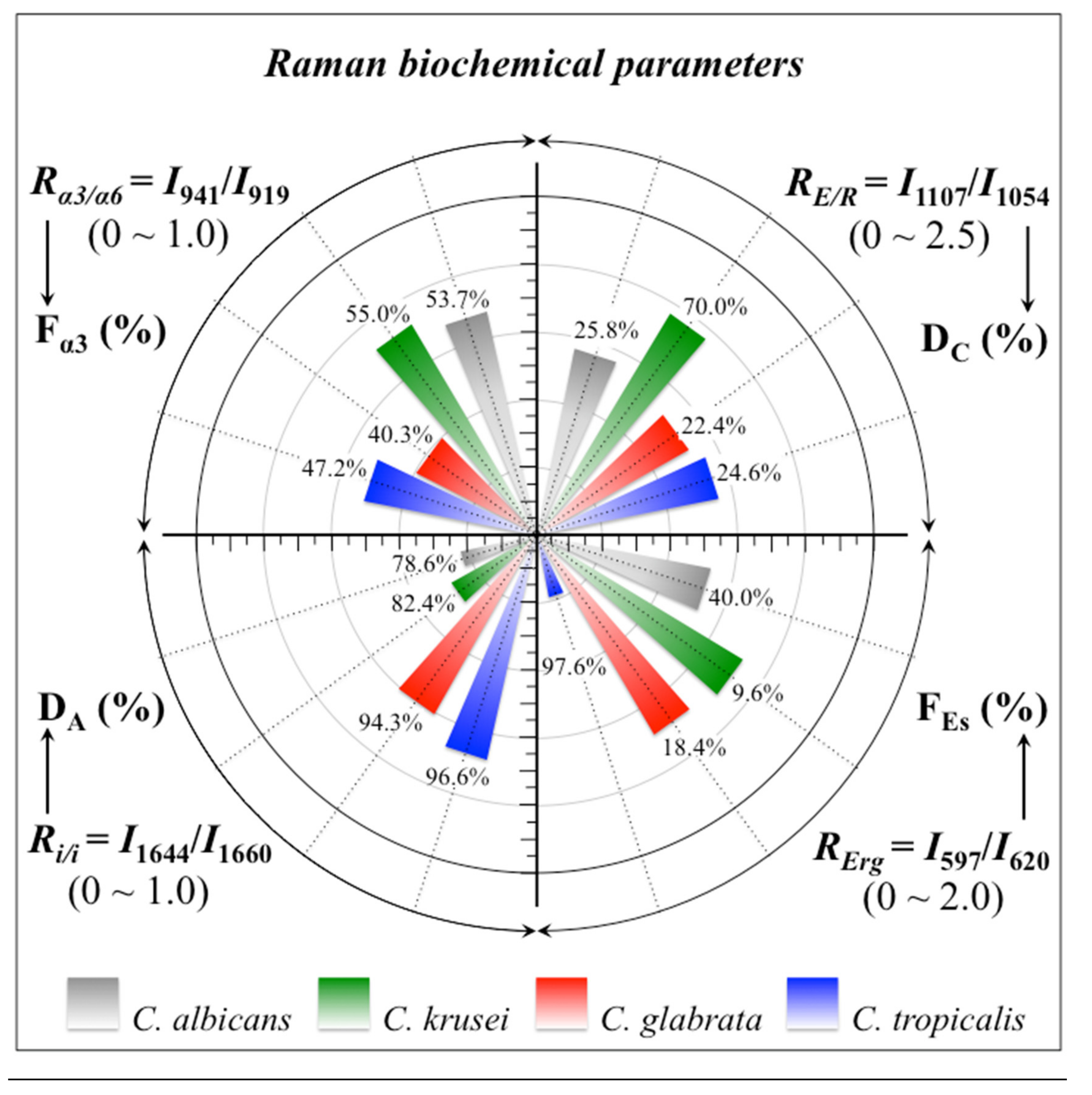
- Polysaccharides—Among Raman bands of polysaccharides, fingerprints for β-1, 3-glucans and α-1, 3-glucans can be found at ~890 cm−1 and ~941 cm−1, respectively. Both bands arise from C-O-C glycosidic linkage vibrations but differ in frequency because of the different bond type and of the structural context in which the bond is embedded. The choice of the 941 cm−1 marker was suggested in previously published studies of α-1, 3-glucans [60,61] Accordingly, the intensity ratio of the 941 to the 890 cm−1 bands, Rα/β = I932/I890, express the volumetric fraction of α- to β-glucans and could be taken as a structural marker for the yeast wall structure. As shown in Figure 3a, the Rα/β ratio clearly differed for different Candida species (i.e., 2.30, 1.76, 1.44, and 2.36 for C. albicans, C. krusei, C. glabrata, and C. tropicalis, respectively). An additional spectroscopic parameter can be envisaged when considering the difference in vibrational behavior between α-1, 3-glucans and α-1, 6-glucans. Figure 4a,b shows the different structures of these two polysaccharides, respectively, and give schematic drafts of their peculiar vibrational fingerprints. As mentioned above, a vibrational signal peculiar to the α-1, 3-glucan structure is the stretching of the C1-O-C3 inter-ring bonds at 941 cm−1 (cf. Figure 4a). On the other hand, vibrational signals peculiar to the α–1, 6–glucan structure can be found at 919 cm−1 (C1-O-C6 inter-ring stretching) and 520 cm−1 (in-plane ring vibration) (cf. Figure 4a). We performed a Raman spectroscopic calibration by mixing α-1, 6-glucan with increasing fractions of α-1, 3-glucan, and recording the respective spectra as shown in Figure 4c. After spectral deconvolution, we computed the Raman intensity ratio, Rα3/α6 = I941/I919, and plotted it as a function of the α-1, 3-glucan fraction, Fα3, in order to obtain a calibration curve as shown in Figure 4d. The availability of a curve linking the Raman parameter, Rα3/α6, to the fraction, Fα3, allows computing the fraction of α-1, 3-glucans in different Candida species. The percent Fα3 values of C. albicans, C. krusei, C. glabrata, and C. tropicalis were calculated from the respective Rα3/α6 values (listed in Figure 3a) as 53.7, 55.0, 40.3, and 47.2%, respectively. The ability of a specific Candida species to synthesize higher fractions of water-insoluble glucans (i.e., glucans rich in α-1,3- and β-1,3-glucosidic linkages) [62], as compared with water-soluble glucans (i.e., rich in α-1,6-glucosidic linkages) [63,64], is crucial in determining the structural and dynamical attributes of the yeast membrane.
- (ii)
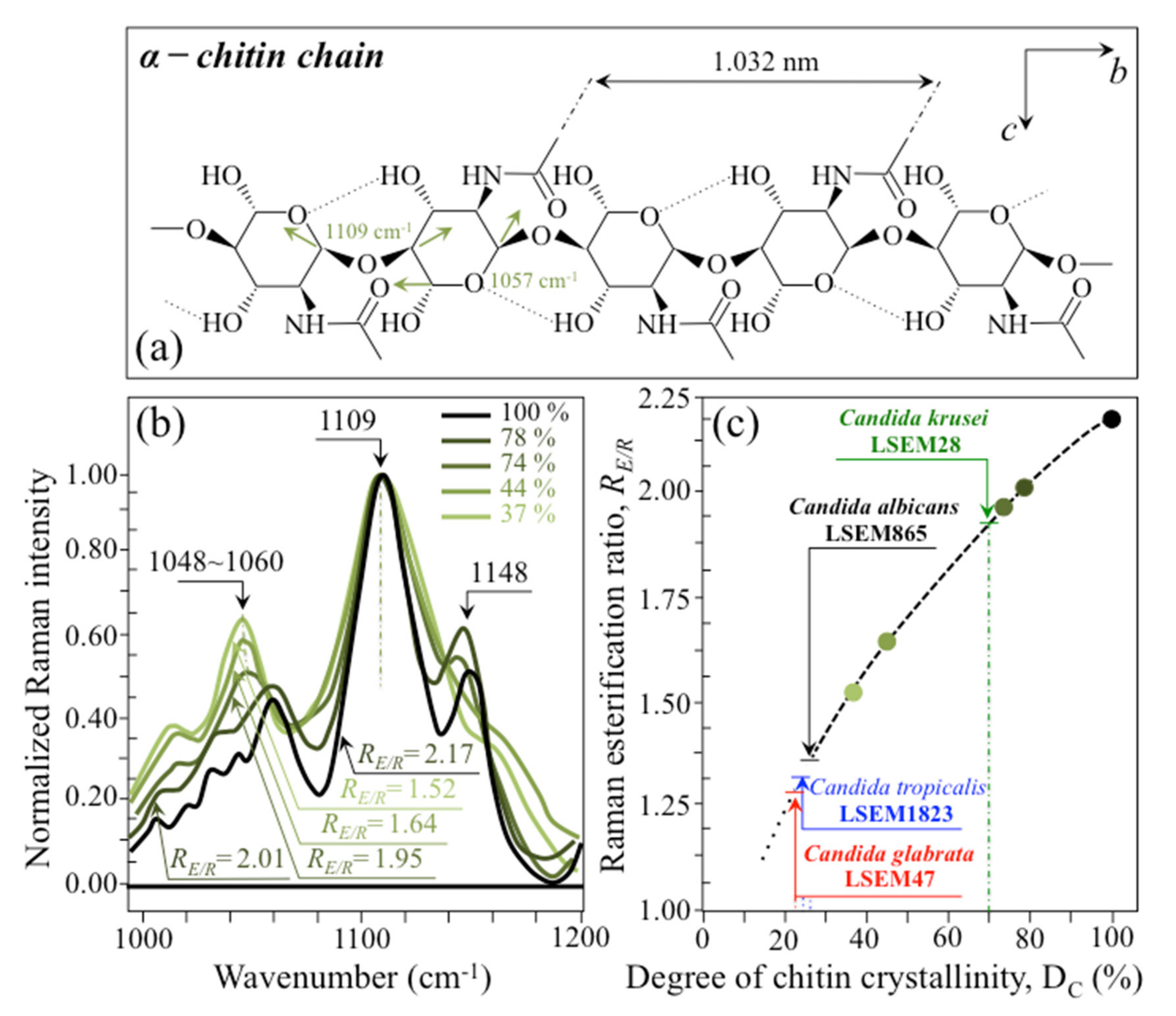
- Chitin backbone—The spectral region representative of C-C-C and C-O-C stretching in polysaccharides corresponds to the wavenumber interval between 950 and 1200 cm−1. Despite structural similarities, which reflect into similar spectroscopic characteristics, distinctions among different polysaccharides can yet be made upon using bands in this peculiar region. Chitin is structurally different from other polysaccharides because of the appearance of C-O-C bonds between neighboring rings and the presence of amide groups to partly replace OH groups as structural sub-units in the polymeric chain [65]. These structural peculiarities confer on chitin distinctive vibrational characteristics as compared to other polysaccharides. The triplet at 1054, 1107, and 1147 cm−1, as well as the low-frequency band at 645 cm−1 stand conspicuously free from overlap with other signals in the spectrum of Candida species, and thus could serve as fingerprints for chitin. On the other hand, as mentioned above, the band at ~1123 cm−1 is a cumulative one for C-C (and C-O) skeletal stretching in polysaccharides. An important detail regarding chitin-related bands lies in the distinction between signals from C-O-C groups within and between rings. The ether C-O-C stretching band between rings is seen at 1107 cm−1, while stretching of C-O-C bonds within rings produces a Raman signal at 1054 cm−1. Accordingly, the band intensity ratio, RE/R = I1107/I1054, can be taken as representative of the ratio between ether and ring C-O-C bonds in chitin. Accordingly, the RE/R ratio could be defined as “esterification ratio”. Figure 5a shows a schematic draft of the chitin chain with its ester and ring vibrational modes. Chitin structures with longer chains are expected to present a higher fraction of ether C-O-C bonds and higher crystallinity; the RE/R ratio thus reflects whether the chitin structure incorporates or not long crystalline chains. C. krusei appears to possess chitin structures with the highest RE/R ratio (1.93), while C. albicans, C. glabrata, and C. tropicalis present RE/R ratios (1.36, 1.27, and 1.30, respectively; cf. labels in inset to Figure 3b). Binias et al. [66] have systematically compared the Raman spectra of natural krill chitin with a series of chitin compounds purposely produced with decreasing chain lengths. These spectra are given in Figure 5b in comparison with the spectrum of a reference (fully crystalline) α-chitin compound (INTIB GmbH, Freiberg, Germany) [67] after normalization to the ether C-O-C stretching signal, seen at the constant wavenumber of 1109 cm−1. The RE/R ratio clearly tended to decrease with decreasing crystallinity (cf. crystallinity values by wide-angle X-ray diffraction measurements in inset to Figure 5b). Note also that unlike the ester C-O-C signal, whose wavenumber remained nearly constant for different degrees of crystallinity, the ring C-O-C signal underwent a clear shift of several wavenumbers (1048~1060 cm−1) with decreasing crystallinity. A plot of the RE/R ratio as a function of percent degree of chitin crystallinity, DC, is drawn in Figure 5c, based on the spectra shown in Figure 5b [66,67,68]. According to this parameter, the chitin structure of C. krusei met the crystalline value closest to standard α-chitin (~70.0%), while C. albicans, C. tropicalis, and C. glabrata presented conspicuously lower DC values (25.8, 24.6, and 22.4%, respectively; cf. Figure 5c). The length of a chitin dimer is 1.023 nm [68] and the average length of a standard α-chitin microfibril is ~33 nm; a chain of fully crystalline α-chitin thus containing on average 64 residues [69]. Assuming the average chain length as proportional to the crystallinity value, it is concluded that, with the exception of C. krusei (average chain length estimated as ~45 residues), C. albicans, C. glabrata, and C. tropicalis incorporate only a small subgroup of crystalline α-chitin allomorph to form rigid microfibrils; whereas the remainders take a different allomorphic structure or just lie in a considerably disordered state due to either unfavorable conformations or unstable hydrogen-bonding patterns [70].
- (iii)
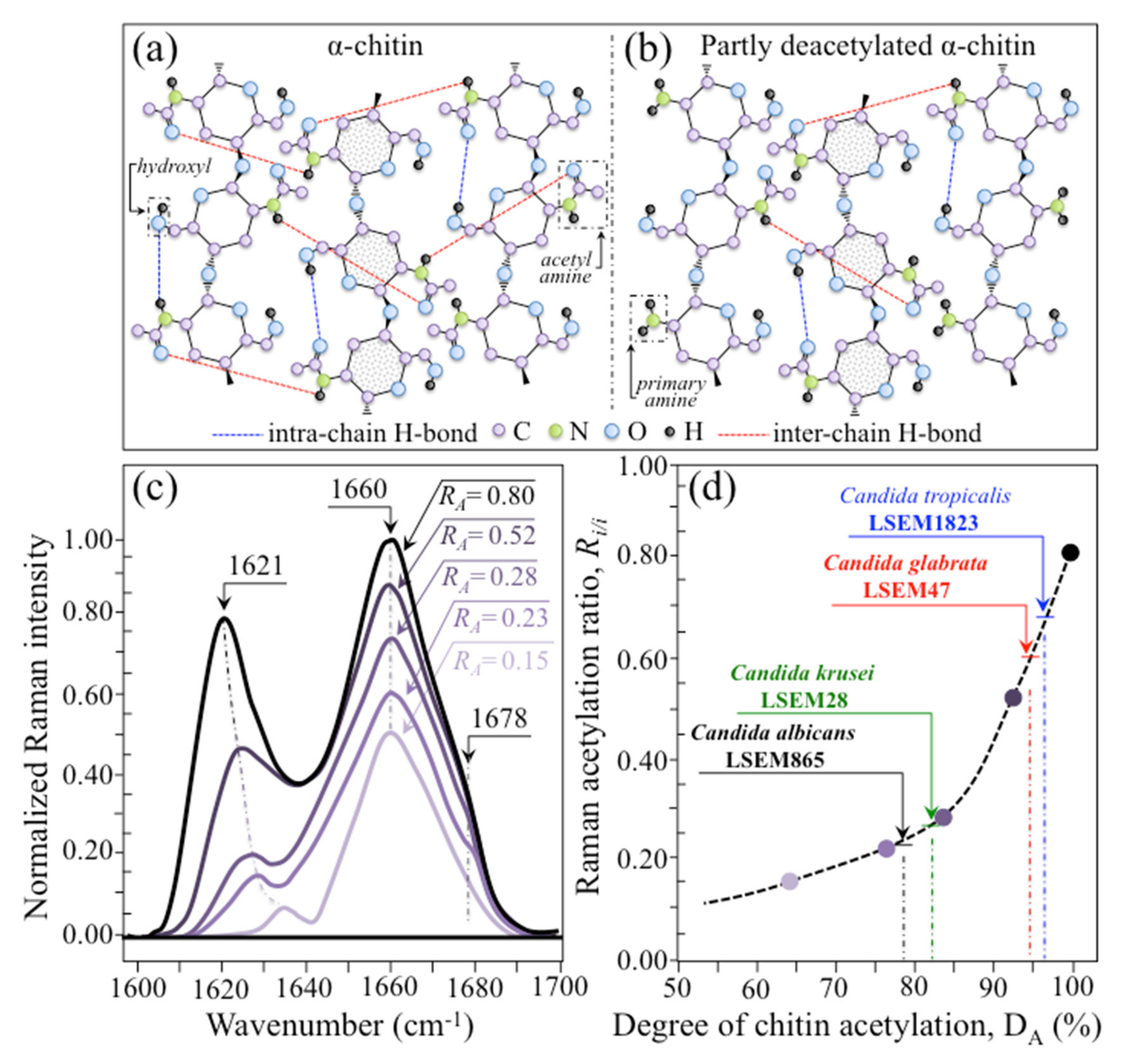
- Chitin sub-units—Amides I, II, and III signals, found in the wavenumber intervals 1600~1700 cm−1, 1400~1500 cm−1, and at around 1300 cm−1, respectively, are peculiar to the N-acetyl group of chitin (cf. labels in Figure 1). The intensities of these signals are thus related to the level of acetylation/deacetylation of the membrane structure of different Candida species. The occurrence of a doublet for the Amide I, which we see at ~1644 and 1660 cm−1 in all species (cf. Figure 3c), has been attributed to the existence of two different types of hydrogen bonds in α-chitin crystals: intermolecular (C=O…HN) and intramolecular (C=O…HO(C6) or C=O…HN) hydrogen bonds (cf. schematic draft in Figure 6a) [71,72,73]. This doublet gives an opportunity to classify different Candida species according to their degree of acetylation. Additional bands in the Amide I region, which are seen as shoulders at 1675~1680 and >1688 cm−1, are assigned to C=O stretching and associated with the presence of different chitin secondary structures or defects in small crystalline chitin fibrils, respectively (cf. Figure 2) [71]. The degree of acetylation, namely, the fraction of acetyl groups present in the α-chitin structure, is linked to the crystalline structure of chitin: a high degree of acetylation favoring the stable α-chitin crystalline structure through a network of hydrogen bonds that the amine acetyl groups can form with NH groups (Figure 6a) [74,75,76]. Conversely, chitins with low degrees of acetylation and, thus, with higher probability of primary amine groups in their structure are more likely to take different allomorphs or more defective structures (cf. Figure 6b). Following Zhang et al. [72], α-chitin structures can be characterized with respect to their degree of acetylation by means of Raman spectroscopy. The Amide I spectra of increasingly deacetylated chitin structures (i.e., containing decreasing fractions of acetyl groups) are shown in Figure 6c. This spectral zone shows three main structure-dependent vibrational characteristics, which vary with decreasing the degree of chitin acetylation, DA: [72] (i) a low-frequency band, seen at ~1621 cm−1 in fully acetylated α-chitin structure, with a decreasing relative intensity and progressively shifting toward higher wavenumbers; (ii) a band located at ~1660 cm−1, decreasing in relative intensity but keeping its spectral location unchanged; and, (iii) a shoulder at 1678 cm−1, showing a decreasing relative intensity. In the Raman spectra shown in Figure 2, the spectral zone between 1602 and 1644 cm−1 did not show Raman signals in any of the studied Candida species; a band seen at 1644 cm−1 was thus assumed as the low-frequency Amide I signal. As mentioned above, one could take the relative intensity ratio, Ri/i = I1644/I1660, referred to as the Raman ratio of intermolecular to intramolecular hydrogen bonds, as a parameter measuring the degree of acetylation of α-chitin. Data from reference chitin structures with known structural characteristics [72] allow building a calibration curve according to which the Ri/i values measured in the Raman spectra of different Candida species can be evaluated and compared (Figure 6d). This procedure reveals that C. tropicalis and C. glabrata possess a relatively high degree of acetylation in their α-chitin structures (i.e., DA = 95~97%), while C. krusei and C. albicans are characterized by relatively more deacetylated α-chitin structures (i.e., DA = 78~82%). Although a high degree of acetylation favors the formation of α-chitin crystalline structure, C. krusei, which showed by far the highest degree of α-chitin crystallization (cf. DC = 70%; cf. Figure 5c) among the studied Candida species, was only the second lowest in degree of acetylation (DA = 82%; cf. Figure 6d); vice versa, C. tropicalis and C. glabrata, both low in DC, were the two highest species in DA. This means that the low DC values of the latter two species are rather due to the presence of different chitin allomorphs than to a disordered state of the α-chitin allomorph itself. As previously mentioned, the structure of α-chitin is stabilized simultaneously by intra- and inter-chain hydrogen bonding (cf. Figure 6a) [71,72,73,77,78]. However, while the former type of hydrogen bond is present in all chitin allomorphs, the latter one is relatively rare in the γ-chitin and completely absent in β-chitin [77,79,80]. Accordingly, C. krusei contains high-crystallinity α-chitin in its most stable, ordered, and tightly packed structure, but it might also incorporate a fraction of β-chitin with no inter-chain bonds, which leads to the relatively low DA measured by Raman spectroscopy. Kaya et al. [81] have measured and compared the Raman spectra of α-, β-, and γ-chitin pure allomorphs. An examination of these spectra revealed that, even on pure compounds, very few spectral details could be available for differentiating different allomorphs from Raman analyses. The machine-learning procedure adopted in this paper located only one relatively hidden but yet resolvable band at ~694 cm−1 that could unequivocally represent the β-chitin allomorph (located with a green asterisk in Figure 2). The relative intensity of this band, which arises from ring deformation [65], is indeed stronger in the spectrum of C. krusei than in different Candida species.
- (iv)
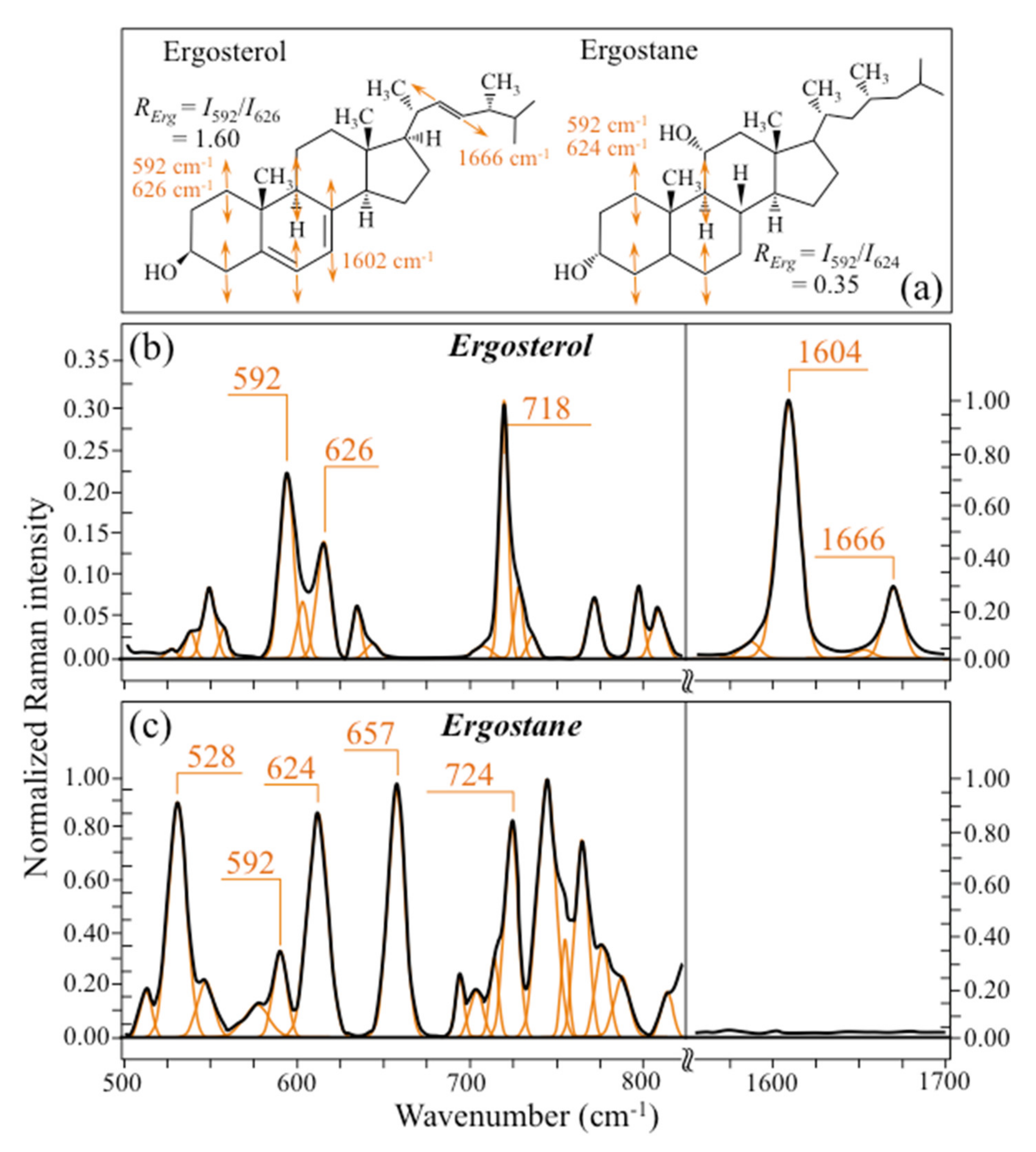
- Ergosterol—Regarding lipids, the machine-learning algorithm located two low-frequency bands (at 594 and 620 cm−1; both related to in-plane ring deformation modes) as mainly (>95%) contributed by steroid ergosterol [82]. Of the two main C=C stretching bands at 1602 cm−1 (within rings) and 1666 cm−1 (in acyl chains), only the former could be detected as a clear shoulder, while the latter undergoes strong overlap with Amide I signals. The relatively intense and isolate band at ~713 cm−1 is contributed by ergosterol (cumulative of C-H bending and ring deformation) [82], but it also contains non-negligible contributions from D-arabitol, chitin, and adenine. Judging from the cumulative (relative) intensities of the 594/620 cm−1 doublet in the spectra of different Candida species (cf. Figure 3d), the fraction of ergosterol appeared to be comparable in all species. However, the trend of ergosterol bands showed important differences in C. tropicalis, as compared with other Candida species:
- (a)
- The doublet at 597/620 cm−1 showed an inversion in relative intensity as compared to the other three Candida species investigated, with the low frequency band becoming more intense than the higher frequency one.
- (b)
- A new band, seen as a shoulder band, was clearly detected at ~650 cm−1, which was not seen in any spectrum of the other Candida species (cf. red asterisk in Figure 2d).
- (c)
- The shoulder band at 1602 cm−1, which was clearly detected in C. albicans, C. krusei, and C. glabrata, conspicuously disappeared in C. tropicalis.
2.3. Raman Imaging of Oral Candida Species
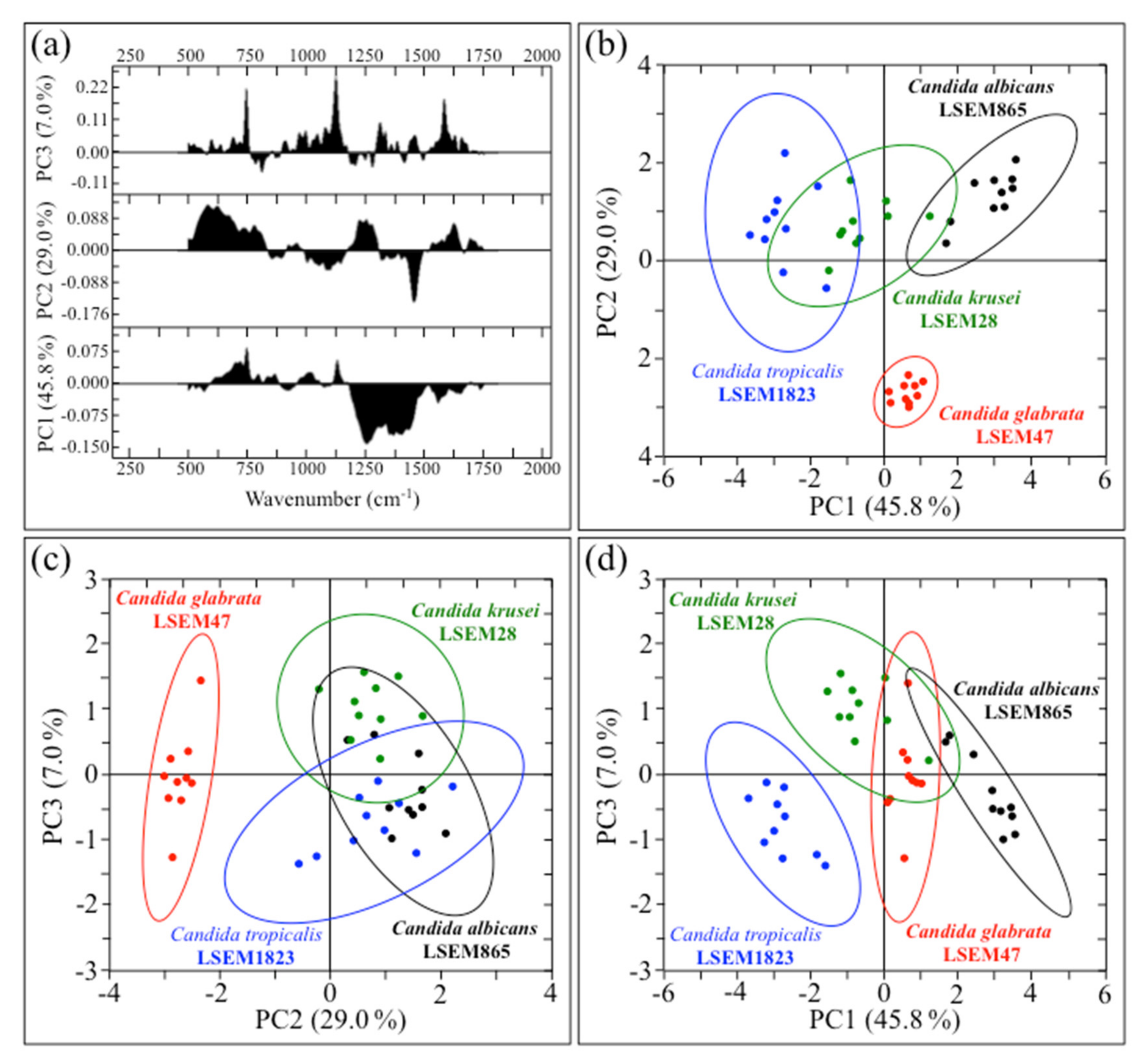
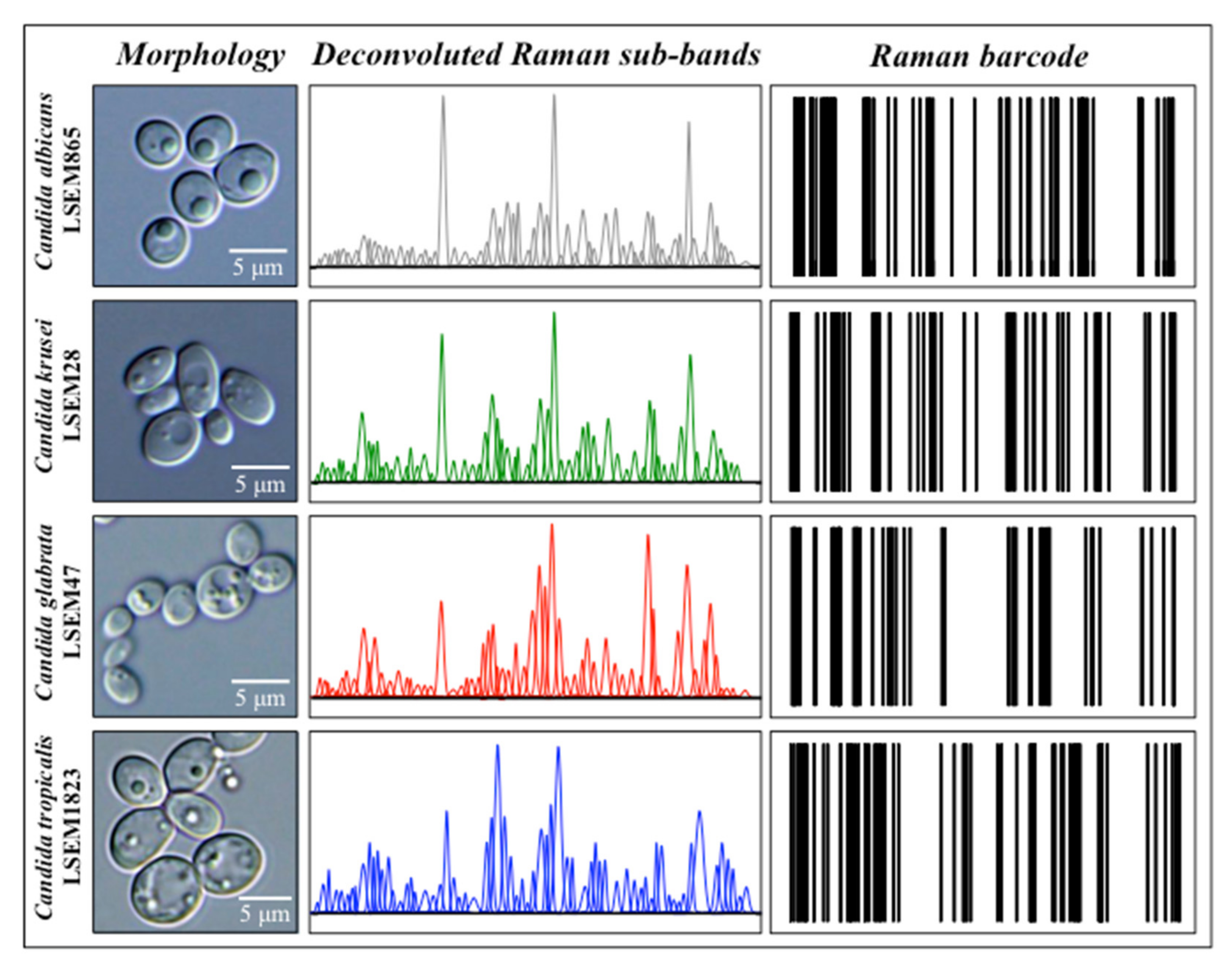
2.4. Chemometric Results and Barcode Construction
3. Discussion
3.1. Interpretation of Raman-Decrypted Molecular-Scale Information
- (i)
- The α-1, 3-glucan is by far the most rigid molecule in the cell-wall structure (followed by chitins and β-glucans), while linear β-1, 3-linked long chains possess the highest level of hydration and mobility [70].
- (ii)
- Chitin and α-1, 3-glucan intertwine to form a rigid and hydrophobic scaffold surrounded by a matrix of pliable and hydrated matrix of β-glucans [70].
- (iii)
- The cell-wall surface is coated with a highly dynamic shell consisting of glycoproteins and a minor fraction of β-glucans [70].
- (iv)
- Non-microfibrillar chitin and β-1, 6-glucan soft chains might link to rigid chitin microfibrils with sufficient lengths to ensure cell-wall flexibility [69].
- (v)
- Different chitin allomorphs coexist as tightly packed chains at the sun-nanometer scale rather than existing in separated longitudinal domains [70].
- (vi)
3.2. Limits of Chemometrics and Leverage of the Barcode Approach
3.3. Clinical Implications of the Proposed Raman Barcode Technology
4. Materials and Methods
4.1. Candida Species
4.2. Differential Interference Contrast (DIC) Microscopy
4.3. In Situ Raman Spectroscopy
4.4. Machine-Learning Algorithm for Spectral Deconvolution
4.5. Chemometric Analysis and Barcoding
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the Oral Fungal Microbiome (Mycobiome) in Healthy Individuals. PLOS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulak-Ozkan, Y.; Kazazoglu, E.; Arikan, A. Oral hygiene habits, denture cleanliness, presence of yeasts and stomatitis in el-derly people. J. Oral Rehabil. 2002, 29, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Wade, W.G. The oral microbiome in health and disease. Pharmacol. Res. 2013, 69, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Oliver, D.E.; Shillitoe, E.J. Effects of smoking on the prevalence and intraoral distribution of Candida albicans. J. Oral Pathol. Med. 1984, 13, 265–270. [Google Scholar] [CrossRef]
- Shimizu, C.; Kuriyama, T.; Williams, D.W.; Karasawa, T.; Inoue, K.; Nakagawa, K.; Yamamoto, E. Association of oral yeast car-riage with specific host factors and altered mouth sensation. Oral Med. 2008, 105, 445–451. [Google Scholar]
- Klein, R.S.; Harris, C.A.; Small, C.B.; Moll, B.; Lesser, M.; Friedland, G.H. Oral Candidiasis in High-Risk Patients as the Initial Manifestation of the Acquired Immunodeficiency Syndrome. N. Engl. J. Med. 1984, 311, 354–358. [Google Scholar] [CrossRef]
- Back-Brito, G.N.; Mota, A.J.; Vasconcellos, T.C.; Querido, S.M.R.; Jorge, A.O.C.; Reis, A.S.M.; Balducci, I.; Koga-Ito, C.Y. Frequency of Candida spp. in the Oral Cavity of Brazilian HIV-Positive Patients and Correlation with CD4 Cell Counts and Viral Load. Mycopathologia 2008, 167, 81–87. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Cheng, H.-H.; Ho, Y.-A.; Hsiao, C.-F.; Lo, H.-J. Fluconazole resistance rate of Candida species from different regions and hospital types in Taiwan. J. Microbiol. Immunol. Infect. 2003, 36, 187–191. [Google Scholar]
- Costa, A.R.; Rodrigues, M.E.; Silva, F.; Henriques, M.; Azeredo, J.; Faustino, A.; Oliveira, R. MIC evaluation of Candida reference strains and clinical isolates by E-test. J. Chemother. 2009, 21, 351–355. [Google Scholar] [CrossRef]
- Negri, M.; Henriques, M.; Svidzinski, T.I.; Paula, R.C.; Oliveira, R. Correlation between Etest, disk diffusion, and micro-dilution methods for antifungal susceptibility testing of Candida species from infection and colonization. J. Clin. Lab. Anal. 2009, 23, 324–330. [Google Scholar] [CrossRef]
- Hunter, K.D.; Gibson, J.; Lockhart, P.; Pithie, A.; Bagg, J. Fluconazole resistant Candida species in the oral flora of fluconazole-exposed HIV-positive patients. Oral Surg. Oral Med. Oral Pathol. 1998, 85, 558–564. [Google Scholar] [CrossRef]
- Martinez, M.; López-Ribot, J.L.; Kirkpatrick, W.R.; Coco, B.J.; Bachmann, S.P.; Patterson, T.F. Replacement of Candida albicans with Candida dubliniensis in human immunodeficiency virus infected patients with oropharyngeal candidiasis treated with fluconazole. J. Clin. Microbiol. 2002, 40, 3135–3139. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A. Antifungal Drug Resistance: Mechanisms, Epidemiology, and Consequences for Treatment. Am. J. Med. 2012, 125 (Suppl. S1), S3–S13. [Google Scholar] [CrossRef]
- Horvath, L.L.; Hospenthal, D.R.; Murray, C.K.; Dooley, D.P. Direct Isolation of Candida spp. from Blood Cultures on the Chromogenic Medium CHROMagar Candida. J. Clin. Microbiol. 2003, 41, 2629–2632. [Google Scholar] [CrossRef] [Green Version]
- Wingard, J.R.; Merz, W.G.; Rinaldi, M.G.; Johnson, T.R.; Karp, J.E.; Saral, R. Increase in Candida krusei Infection among Patients with Bone Marrow Transplantation and Neutropenia Treated Prophylactically with Fluconazole. N. Engl. J. Med. 1991, 325, 1274–1277. [Google Scholar] [CrossRef]
- Abbas, J.; Bodey, G.P.; Hanna, H.A.; Mardani, M.; Girgawy, E.; Abi-Said, D.; Whimbey, E.; Hachem, R.; Raab, I. Candida krusei fungemia: An escalating serious infection in immunocompromised patients. Arch. Intern. Med. 2000, 160, 2659–2664. [Google Scholar] [CrossRef] [Green Version]
- Berrouane, Y.F.; Hollis, R.J.; Pfaller, M.A. Strain variation among and antifungal susceptibilities of isolates of Candida krusei. J. Clin. Microbiol. 1996, 34, 1856–1858. [Google Scholar] [CrossRef] [Green Version]
- Hakki, M.; Staab, J.F.; Marr, K.A. Emergence of a Candida krusei Isolate with Reduced Susceptibility to Caspofungin during Therapy. Antimicrob. Agents Chemother. 2006, 50, 2522–2524. [Google Scholar] [CrossRef] [Green Version]
- Kahn, J.N.; Garcia-Effron, G.; Hsu, M.J.; Park, S.; Marr, K.A.; Perlin, D.S. Acquired echinocandin resistance in a Candida krusei isolate due to modification of glucan synthase. Antimicrob. Agents Chemother. 2007, 51, 1876–1878. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of Invasive Candidiasis: A Persistent Public Health Problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Takata, T.; Akamatsu, S.; Hatakenaka, K.; Matsumoto, S.; Maki, K.; Ikeda, F.; Tamai, R.; Tamura, K. Analysis of Candida glabrata Strains with Reduced Sensitivity to Micafungin In Vitro Isolated from a Patient with Persistent Candidemia. Jpn. J. Infect. Dis. 2010, 63, 332–337. [Google Scholar] [CrossRef]
- Pham, C.D.; Iqbal, N.; Bolden, C.B.; Kuykendall, R.J.; Harrison, L.H.; Farley, M.M.; Schaffner, W.; Beldavs, Z.G.; Chiller, T.M.; Park, B.J.; et al. Role of FKS Mutations in Candida glabrata: MIC Values, Echinocandin Resistance, and Multidrug Resistance. Antimicrob. Agents Chemother. 2014, 58, 4690–4696. [Google Scholar] [CrossRef] [Green Version]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Kwa, A.L.; Cheng, S.; Du, C.; Clancy, C.J. The Presence of an FKS Mutation Rather than MIC Is an Independent Risk Factor for Failure of Echinocandin Therapy among Patients with Invasive Candidiasis Due to Candida glabrata. Antimicrob. Agents Chemother. 2012, 56, 4862–4869. [Google Scholar] [CrossRef] [Green Version]
- Bizerra, F.C.; Jimenez-Ortigosa, C.; Souza, A.C.; Breda, G.L.; Queiroz-Telles, F.; Perlin, D.S.; Colombo, A.L. Breakthrough candidemia due to multidrug resistant C. glabrata during prophylaxis with low dose of micafungin. Antimicrob. Agents Chemother. 2014, 58, 2438–2440. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.-H.; Jung, D.S.; Lee, J.Y.; Kim, H.A.; Ryu, S.; Jung, S.-I.; Joo, E.-J.; Cheon, S.; Kim, Y.-S.; Kim, S.-W.; et al. Poor prognosis of Candida tropicalis among non-albicans candidemia: A retrospective multi-center cohort study, Korea, Diagn. Microbiol. Infect. Dis. 2019, 95, 195–200. [Google Scholar]
- Arendrup, M.C.; Bruun, B.; Christensen, J.J.; Fuursted, K.; Johansen, H.K.; Kjældgaard, P.; Knudsen, J.D.; Kristensen, L.; Møller, J.; Nielsen, L.; et al. National surveillance of fungemia in Denmark, (2004 to 2009). J. Clin. Microbiol. 2011, 49, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.-Y.; Chuang, Y.-C.; Wu, U.-I.; Sun, H.-Y.; Wang, J.-T.; Sheng, W.-H.; Lo, H.-J.; Wang, H.-Y.; Chen, Y.-C.; Chang, S.-C. Clonality of fluconazole-nonsusceptible Candida tropicalis in bloodstream infections, Taiwan, 2011–2017. Emerg. Infect. Dis. 2019, 25, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Megri, Y.; Arastehfar, A.; Boekhout, T.; Daneshnia, F.; Hörtnagl, C.; Sartori, B.; Hafez, A.; Pan, W.; Lass-Flörl, C.; Hamrioui, B. Candida tropicalis is the most prevalent yeast species causing candidemia in Algeria: The urgent need for antifungal stew-ardship and infection control measures. Antimicrob. Resist. Infect. Control 2020, 9, 50. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty years of the SENTRY antifungal surveillance program: Results for Candida species from 1997–2016. Open Forum Infect. Dis. 2019, 6 (Suppl. S1), S79–S94. [Google Scholar] [CrossRef] [Green Version]
- Arthington-Skaggs, B.A.; Lee-Yang, W.; Ciblak, M.A.; Frade, J.P.; Brandt, M.E.; Hajjeh, R.A.; Harrison, L.H.; Sofair, A.N.; Warnock, A.D.W. Comparison of Visual and Spectrophotometric Methods of Broth Microdilution MIC End Point Determination and Evaluation of a Sterol Quantitation Method for In Vitro Susceptibility Testing of Fluconazole and Itraconazole against Trailing and Nontrailing Candida Isolates. Antimicrob. Agents Chemother. 2002, 46, 2477–2481. [Google Scholar] [CrossRef] [Green Version]
- Rueda, C.; Puig-Asensio, M.; Guinea, J.; Almirante, B.; Cuenca-Estrella, M.; Zaragoza, O.; Padilla, B.; Muñoz, P.; Pardo, J.P.; García-Rodríguez, J.; et al. Evaluation of the possible influence of trailing and paradoxical effects on the clinical outcome of patients with candidemia. Clin. Microbiol. Infect. 2016, 23, 49.e1–49.e8. [Google Scholar] [CrossRef] [Green Version]
- Pezzotti, G.; Kobara, M.; Asai, T.; Nakaya, T.; Miyamoto, N.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Ohgitani, E.; Marin, E.; et al. Raman Imaging of Pathogenic Candida auris: Visualization of Structural Characteristics and Machine-Learning Identification. Front. Microbiol. 2021, 12, 769597. [Google Scholar] [CrossRef]
- Pezzotti, G.; Kobara, M.; Nakaya, T.; Asai, T.; Miyamoto, N.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Ohgitani, E.; Marin, E.; et al. Raman study of pathogenic Candida auris: Imaging metabolic machineries in reaction to antifungal drugs. Front. Microbiol. in press.
- Pezzotti, G.; Boschetto, F.; Ohgitani, E.; Fujita, Y.; Shin-Ya, M.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Marin, E.; Zhu, W.; et al. Raman molecular fingerprints of SARS-CoV-2 British variant and the concept of Raman barcode. Adv. Sci. 2021, 2021, 2103287. [Google Scholar] [CrossRef] [PubMed]
- Pezzotti, G. Raman spectroscopy in cell biology and microbiology. J. Raman Spectrosc. 2021, 52, 2348–2443. [Google Scholar] [CrossRef]
- Potocki, L.; Depciuch, J.; Kuna, E.; Worek, M.; Lewinska, A.; Wnuk, M. FTIR and Raman spectroscopy-based biochemical profiling reflects genomic diversity of clinical Candida isolates that may be useful for diagnosis and targeted therapy of candidiasis. Int. J. Mol. Sci. 2019, 20, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samek, O.; Mlynarikova, K.; Bernatova, S.; Ježek, J.; Krzyzánek, V.; Siler, M.; Zemanek, P.; Ruzicka, F.; Holá, V.; Mahelová, M. Candida parapsilosis Biofilm Identification by Raman Spectroscopy. Int. J. Mol. Sci. 2014, 15, 23924–23935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebrošová, K.; Šiler, M.; Samek, O.; Růžička, F.; Bernatová, S.; Ježek, J.; Zemánek, P.; Holá, V. Identification of ability to form biofilm in Candida parapsilosis and Staphylococcus epidermidis by Raman spectroscopy. Futur. Microbiol. 2019, 14, 509–517. [Google Scholar] [CrossRef]
- Chouthai, N.S.; Shah, A.A.; Salimnia, H.; Palyvoda, O.; Devpura, S.; Klein, M.; Asmar, B. Use of Raman Spectroscopy to Decrease Time for Identifying the Species of Candida Growth in Cultures. Avicenna J. Med. Biotechnol. 2015, 7, 45–48. [Google Scholar]
- Puanglek, S.; Kimura, S.; Enomoto-Rogers, Y.; Kabe, T.; Yoshida, M.; Wada, M.; Iwata, T. In vitro synthesis of linear α-1,3-glucan and chemical modification to ester derivatives exhibiting outstanding thermal properties. Sci. Rep. 2016, 6, 30479. [Google Scholar] [CrossRef] [PubMed]
- Brunke, S.; Hube, B. Two unlike cousins: Candida albicans and C. glabrata infection strategies. Cell. Microbiol. 2013, 15, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Sachivkina, N.; Podoprigora, I.; Bokov, D. Morphological characteristics of Candida albicans, Candida krusei, Candida guilliermondii, and Candida glabrata biofilms, and response to farnesol. Vet. World 2021, 14, 1608–1614. [Google Scholar] [CrossRef]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; de Montigny, J.; Marck, C.; Neuveglise, C.; Goffard, N.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef]
- Huang, Z.; McWilliams, A.; Lui, H.; McLean, D.I.; Lam, S.; Zeng, H. Near-infrared Raman spectroscopy for optical diagnosis of lung cancer. Int. J. Cancer 2003, 107, 1047–1052. [Google Scholar] [CrossRef]
- Zhu, G.; Zhu, X.; Fan, Q.; Wan, X. Raman spectra of amino acids and their aqueous solutions. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2010, 78, 1187–1195. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, J.; Tian, L.; Zhang, Q.; Guan, Y.; Chen, L.; Liu, G.; Yu, H.-Q.; Tian, Y.; Huang, Q. Raman micro-spectroscopy monitoring of cytochrome c redox state in Candida utilis during cell death under low-temperature plasma-induced oxidative stress. Analyst 2020, 145, 3922–3930. [Google Scholar] [CrossRef]
- Welchen, E.; Gonzalez, D.H. Cytochrome c, a hub linking energy, redox, stress and signaling pathways in mitochondria and other cell compartments. Physiol. Plant. 2016, 157, 310–321. [Google Scholar] [CrossRef]
- Alvarez-Paggi, D.; Hannibal, L.; Castro, M.A.; Oviedo-Rouco, S.; Demicheli, V.; Tórtora, V.; Tomasina, F.; Radi, R.; Murgida, D.H. Multifunctional Cytochrome c: Learning New Tricks from an Old Dog. Chem. Rev. 2017, 117, 13382–13460. [Google Scholar] [CrossRef]
- Okada, M.; Smith, N.I.; Palonpon, A.F.; Endo, H.; Kawata, S.; Sodeoka, M.; Fujita, K. Label-free Raman observation of cytochrome c dynamics during apoptosis. Proc. Natl. Acad. Sci. USA 2012, 109, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.C.; Borutaite, V. Regulation of apoptosis by the redox state of cytochrome c. Biochim. Biophys. Acta (BBA) Bioenerg. 2008, 1777, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Kakita, M.; Kaliaperumal, V.; Hamaguchi, H. Resonance Raman quantification of the redox state of cytochromes b and c in-vivo and in-vitro. J. Biophoton. 2012, 5, 20–24. [Google Scholar] [CrossRef]
- Hüttemann, M.; Pecina, P.; Rainbolt, M.; Sanderson, T.H.; Kagan, V.E.; Samavati, L.; Doan, J.W.; Lee, I. The multiple functions of cytochrome c and their regulation in life and death decisions of the mammalian cell: From respiration to apoptosis. Mitochondrion 2011, 11, 369–381. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, F.; Cammisuli, F.; Addobbati, R.; Rizzardi, C.; Gessini, A.; Masciovecchio, C.; Rossi, B.; Pascolo, L. Oxidative damage in DNA base revealed by UV resonant Raman spectroscopy. Analyst 2015, 140, 1477–1485. [Google Scholar] [CrossRef]
- Barboza, F.; Filho, J.D.S.; Freire, P.; Filho, P.F.; Lima, J.; Melo, F.; Joya, M.; Barba-Ortega, J. High-pressure Raman spectra of thymidine crystals. Vib. Spectrosc. 2017, 89, 62–68. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Wickes, B.L.; Whelan, W.L. Ploidy and DNA content of Candida stelloidea cells. Infect. Immun. 1987, 55, 3207–3208. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.S.; Magee, B.B.; Sheppard, D.C.; Yang, M.; Kauffman, S.; Becker, J.; Edwards, J.E., Jr.; Magee, P.T. Effects of ploidy and mating type on virulence of Candida albicans. Infect. Immun. 1987, 73, 7366–7374. [Google Scholar] [CrossRef] [Green Version]
- Seervai, R.N.H.; Jones, S.K., Jr.; Hirakawa, M.P.; Porman, A.M.; Bennett, R.J. Parasexuality and ploidy change in Candida tropicalis. Eukaryot. Cell 2013, 12, 1629–1640. [Google Scholar] [CrossRef] [Green Version]
- Berman, J. Ploidy plasticity: A rapid and reversible strategy for adaptation to stress. FEMS Yeast Res. 2016, 16, fow020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, C.; Fraser, J. Ploidy variation as an adaptive mechanism in human pathogenic fungi. Semin. Cell Dev. Biol. 2013, 24, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spevacek, J.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Mikkelsen, M.S.; Jespersen, B.M.; Møller, B.L.; Lærke, H.N.; Larsen, F.H.; Engelsen, S.B. Comparative spectroscopic and rheological studies on crude and purified soluble barley and oat β-glucan preparations. Food Res. Int. 2010, 43, 2417–2424. [Google Scholar] [CrossRef]
- Hanada, N.; Kuramitsu, H.K. Isolation and characterization of the Streptococcus mutans gtfC gene, coding for synthesis of both soluble and insoluble glucans. Infect. Immun. 1988, 56, 1999–2005. [Google Scholar] [CrossRef] [Green Version]
- Hanada, N.; Kuramitsu, H.K. Isolation and characterization of the Streptococcus mutans gtfD gene, coding for primer-dependent soluble glucan synthesis. Infect. Immun. 1989, 57, 2079–2085. [Google Scholar] [CrossRef] [Green Version]
- Wangpaiboon, K.; Padungros, P.; Nakapong, S.; Charoenwongpaiboon, T.; Rejzek, M.; Field, R.; Pichyangkura, R. An α-1,6-and α-1,3-linked glucan produced by Leuconostoc citreum ABK-1 alternansucrase with nanoparticle and film-forming properties. Sci. Rep. 2018, 8, 8340. [Google Scholar] [CrossRef]
- De Gussem, K.; Vandenabeele, P.; Verbeken, A.; Moens, L. Raman spectroscopic study of Lactarius spores (Russulales, Fungi). Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2005, 61, 2896–2908. [Google Scholar] [CrossRef]
- Binias, D.; Wyszomirski, M.; Binias, W.; Boryniec, S. Supermolecular structure of chitin and its derivatives in FTIR spectroscopy studies. Prog. Chem. Appl. Chitin Its Deriv. 2007, XII, 95–108. [Google Scholar]
- Ehrlich, H.; Shaala, L.A.; Youssef, D.T.A.; Żółtowska- Aksamitowska, S.; Tsurkan, M.; Galli, R.; Meissner, H.; Wysokowski, M.; Petrenko, I.; Tabachnick, K.R.; et al. Discovery of chitin in skeletons of non-verongiid Red Sea demosponges. PLoS ONE 2018, 13, e0195803. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.F. Arthropod cuticle: A natural composite shell system. Compos. Part A Appl. Sci. Manuf. 2002, 33, 1311–1315. [Google Scholar] [CrossRef]
- Lenardon, M.D.; Sood, P.; Dorfmueller, H.C.; Brown, A.J.; Gow, N.A. Scalar nanostructure of the Candida albicans cell wall; a molecular, cellular and ultrastructural analysis and interpretation. Cell Surf. 2020, 6, 100047. [Google Scholar] [CrossRef]
- Kang, X.; Kirui, A.; Muszyński, A.; Widanage, M.C.D.; Chen, A.; Azadi, P.; Wang, P.; Mentink-Vigier, F.; Wang, T. Molecular architecture of fungal cell walls revealed by solid-state NMR. Nat. Commun. 2018, 9, 2747. [Google Scholar] [CrossRef] [Green Version]
- Focher, B.; Naggi, A.; Torri, G.; Cosani, A.; Terbojevich, M. Structural differences between chitin polymorphs and their precipi-tates from solutions—Evidence from CP-MAS13C-NMR, FT-IR and FT-Raman spectroscopy. Carbohydr. Polym. 1992, 17, 97–102. [Google Scholar] [CrossRef]
- Zhang, K.; Geissler, A.; Fischer, S.; Brendler, E.; Baucker, E. Solid-state spectroscopic characterization of α-chitins deacetylated in homogeneous solutions. J. Phys. Chem. B 2012, 116, 4584–4592. [Google Scholar] [CrossRef]
- Minke, R.; Blackwell, J. The structure of α–Chitin. J. Mol. Biol. 1978, 120, 167–181. [Google Scholar] [CrossRef]
- Wu, Y.; Sasaki, T.; Irie, S.; Sakurai, K. A novel biomass-ionic liquid platform for the utilization of native chitin. Polymer 2008, 49, 2321–2327. [Google Scholar] [CrossRef]
- Okuyama, K.; Noguchi, A.K.; Miyazawa, T.; Yui, T.; Ogawa, K. Molecular and Crystal Structure of Hydrated Chitosan. Macromolecules 1997, 30, 5849–5855. [Google Scholar] [CrossRef]
- Okuyama, K.; Noguchi, K.; Kanenari, M.; Egawa, T.; Osawa, K.; Ogawa, K. Structural diversity of chitosan and its complexes. Carbohydr. Polym. 2000, 41, 237–247. [Google Scholar] [CrossRef]
- Kameda, T.; Miyazawa, M.; Ono, H.; Yoshida, M. Hydrogen bonding structure and stability of -chitin studied by 13C Solid-State NMR. Macromol. Biosci. 2005, 5, 103–106. [Google Scholar] [CrossRef]
- Deringer, V.L.; Englert, U.; Dronskowski, R. Nature, strength, and cooperativity of the hydrogen-bonding network in α-chitin. Biomacromolecules 2016, 17, 996–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, D.; Nishiyama, Y.; Langan, P.; Forsyth, V.T.; Kimura, S.; Wada, M. Water in crystalline fibers of dehydrate β-chitin results in unexpected absence of intramolecular hydrogen bonding. PLoS ONE 2012, 7, e39376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, D.; Nishiyama, Y.; Langan, P.; Forsyth, V.T.; Kimura, S.; Wada, M. Direct Determination of the Hydrogen Bonding Arrangement in Anhydrous β-Chitin by Neutron Fiber Diffraction. Biomacromolecules 2012, 13, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Mujtaba, M.; Ehrich, H.; Salaberria, A.; Baran, T.; Amemiya, C.; Galli, R.; Akyuz, L.; Sargin, I.; Labidi, J. On chemistry of γ-chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Živanović, V.; Semini, G.; Laue, M.; Drescher, D.; Aebischer, T.; Kneipp, J. Chemical Mapping of Leishmania Infection in Live Cells by SERS Microscopy. Anal. Chem. 2018, 90, 8154–8161. [Google Scholar] [CrossRef]
- Edwards, H.G.M.; Herschy, B.; Page, K.; Munshi, T.; Scowen, I.J. Raman spectra of biomarkers of relevance to analytical as-trobiological exploration: Hopanoids, sterols and steranes. Spectrochim. Acta Part A 2011, 78, 191–195. [Google Scholar] [CrossRef]
- Munro, C.A. Chitin and Glucan, the Yin and Yang of the Fungal Cell Wall, Implications for Antifungal Drug Discovery and Therapy. Adv. Appl. Microbiol. 2013, 83, 145–172. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden Killers: Human Fungal Infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [Green Version]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Barreto-Bergter, E.B.; Figueiredo, R.T. Fungal glycans and the innate immune recognition. Front. Cell. Infect. Microbiol. 2014, 4, 145. [Google Scholar] [CrossRef] [Green Version]
- Rappleye, C.A.; Eissenberg, L.G.; Goldman, W.E. Histoplasma capsulatum alpha-(1,3)-glucan blocks innate immune recognition by the beta-glucan receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 1366–1370. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Domergue, R.; Zupancic, M.L.; Cormack, B.P. A yeast by any other name: Candida glabrata and its interaction with the host. Curr. Opin. Microbiol. 2005, 8, 378–384. [Google Scholar] [CrossRef]
- Kaloriti, D.; Tillmann, A.; Cook, E.; Jacobsen, M.; You, T.; Lenardon, M.; Ames, L.; Barahona, M.; Chandrasekaran, K.; Coghill, G.; et al. Combinatorial stresses kill pathogenic Candida species. Med. Mycol. 2012, 50, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Roetzer, A.; Gregori, C.; Jennings, A.M.; Quintin, J.; Ferrandon, D.; Butler, G.; Kuchler, K.; Ammerer, G.; Schueller, C. Candida glabrata environmental stress response involves Saccharomyces cerevisiae Msn2/4 orthologous transcription factors. Mol. Microbiol. 2008, 69, 603–620. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Yao, S. The effect of created local hyperosmotic microenvironment in microcapsule for the growth and metabolism of osmotolerant yeast Candida krusei. BioMed Res. Int. 2013, 2013, 467263. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-J.; Liu, D.-H.; Zhong, J.-J. Novel Fermentation Strategy for Enhancing Glycerol Production by Candida krusei. Biotechnol. Prog. 2003, 19, 1615–1619. [Google Scholar] [CrossRef]
- Dang, H.V.; Marini, F. Editorial: Chemometrics-based Spectroscopy for Pharmaceutical and Biomedical Analysis. Front. Chem. 2019, 7, 153. [Google Scholar] [CrossRef]
- Gautam, R.; Vanga, S.; Ariese, F.; Umapathy, S. Review of multidimensional data processing approaches for Raman and infrared spectroscopy. EPJ Tech. Instrum. 2015, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Deming, S.N. Chemometrics: An overview. Clin. Chem. 1986, 32, 1702–1706. [Google Scholar] [CrossRef]
- Koppel, R.; Wetterneck, T.; Telles, J.L.; Karsh, B.-T. Workarounds to barcode medication administration systems: Their occurrences, causes, and threats to patient safety. J. Am. Med. Inform. Assoc. 2008, 15, 408–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebonah, D.E.; Dileep, A.; Chandrasekhar, K.; Sreevani, S.; Sreedevi, B.; Kumari, J.P. DNA Barcoding on Bacteria: A Review. Adv. Biol. 2014, 2014, 541787. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.J.M.; Crous, P.W.; Boekhout, T.; et al. DNA barcoding analysis of more than 9000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Siddall, M.E.; Fontanella, F.M.; Watson, S.C.; Kvist, S.; Erséus, C. Barcoding Bamboozled by Bacteria: Convergence to Metazoan Mitochondrial Primer Targets by Marine Microbes. Syst. Biol. 2009, 58, 445–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savolainen, V.; Cowan, R.S.; Vogler, A.P.; Roderick, G.K.; Lane, R. Towards writing the encyclopaedia of life: An introduction to DNA barcoding, Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1805–1811. [Google Scholar] [CrossRef]
- Forastiero, A.; Garcia-Gil, V.; Rivero-Menendez, O.; Garcia-Rubio, R.; Monteiro, M.C.; Alastruey-Izquierdo, A.; Jordan, R.; Agorio, I.; Mellado, E. Rapid Development of Candida krusei Echinocandin Resistance during Caspofungin Therapy. Antimicrob. Agents Chemother. 2015, 59, 6975–6982. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pezzotti, G.; Kobara, M.; Nakaya, T.; Imamura, H.; Miyamoto, N.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Ohgitani, E.; Marin, E.; et al. Raman Spectroscopy of Oral Candida Species: Molecular-Scale Analyses, Chemometrics, and Barcode Identification. Int. J. Mol. Sci. 2022, 23, 5359. https://doi.org/10.3390/ijms23105359
Pezzotti G, Kobara M, Nakaya T, Imamura H, Miyamoto N, Adachi T, Yamamoto T, Kanamura N, Ohgitani E, Marin E, et al. Raman Spectroscopy of Oral Candida Species: Molecular-Scale Analyses, Chemometrics, and Barcode Identification. International Journal of Molecular Sciences. 2022; 23(10):5359. https://doi.org/10.3390/ijms23105359
Chicago/Turabian StylePezzotti, Giuseppe, Miyuki Kobara, Tamaki Nakaya, Hayata Imamura, Nao Miyamoto, Tetsuya Adachi, Toshiro Yamamoto, Narisato Kanamura, Eriko Ohgitani, Elia Marin, and et al. 2022. "Raman Spectroscopy of Oral Candida Species: Molecular-Scale Analyses, Chemometrics, and Barcode Identification" International Journal of Molecular Sciences 23, no. 10: 5359. https://doi.org/10.3390/ijms23105359